| |||
| |||
Συναπτική Πλαστικότητα Ι: Ενδείξεις για την ηλεκτροδομική
βάση της Μακρόχρονης Αποδυνάμωσης (LTD)
ΜΙΧΜΙΖΟΣ Δ., ΚΩΣΤΑ Β., ΚΑΡΛΟΒΑΣΙΤΟΥ-ΚΟΝΙΑΡΗ Α., ΑΣΠΡΟΔΙΝΗ Ε., ΜΠΑΛΟΓΙΑΝΝΗΣ
Σ.
Α’ Νευρολογική Κλινική, ΑΠΘ, ΑΧΕΠΑ
Περίληψη
Από την πρώτη παρατήρησή της, από τους Bliss και Lψmo το 1973, η συναπτική
πλαστικότητα θεωρούνταν ως το πιθανότερο πειραματικό παράδειγμα να μας προσφέρει
την κατανόηση του πως αποθηκεύονται οι πληροφορίες στον εγκέφαλο των σπονδυλωτών.
Τα τελευταία 30 χρόνια έχουν αποδειχθεί διάφορες μορφές συναπτικής πλαστικότητας,
με πλέον σημαντικές τη μακρόχρονη ενδυνάμωση (LTP) και τη μακρόχρονη αποδυνάμωση
(LTD), και τα χαρακτηριστικά και οι απαιτούμενες συνθήκες πρόκλησής τους είναι
πολύ ενδεικτικά του ότι αυτή η πλαστικότητα αποτελεί το υπόστρωμα της μνημονικής
εγχάραξης. Πειραματική πρόκληση σταθερής LTD στην CA1 περιοχή του ιππόκαμπου
και μετέπειτα παρατήρηση των δομικών μετατροπών των συνάψεων στο ηλεκτρονικό
μικροσκόπιο αποκάλυψε ότι η συναπτική αποδυνάμωση είναι το αποτέλεσμα ηλεκτροδομικών
μετατροπών των ακανθικών συνάψεων. Προτείνεται ότι το ερέθισμα χαμηλόσυχνης
ριπής, που επιφέρει LTD, προκαλεί μία περιορισμένη εισροή ιόντων Ca++ με την
αύξηση όμως της συγκέντρωσής τους να περιορίζεται στην περιμεμβρανική περιοχή
της σύναψης. Αυτή η αύξηση επιφέρει κυτταροσκελετικές μετατροπές οι οποίες έχουν
ως αποτέλεσμα την "διάτρηση" της μετασυναπτικής πύκνωσης (PSD) και την αύξηση
της μεμβρανικής επιφάνειας. Καθώς όμως η αύξηση της συγκέντρωσης ιόντων Ca++
είναι μόνο σύντομη και παροδική, δεν επαρκεί για την απορρόφηση υποδοχέων και
ιοντικών διαύλων από το δενδριτικό στέλεχος, ώστε να καταστεί λειτουργική η
αυξημένη μεμβρανική επιφάνεια, και έτσι η αυξημένη επιφάνεια της μετασυναπτικής
μεμβράνης παρουσιάζει διογκωμένη ηλεκτρική αντίσταση.
Λέξεις κλειδιά: Μνήμη, ιππόκαμπος, συναπτική πλαστικότητα, μακρόχρονη αποδυνάμωση (LTD).
Εισαγωγή
Η ιδέα της αποθήκευσης πληροφοριών στον εγκέφαλο με τη μορφή αλλαγών της συναπτικής αποτελεσματικότητας έχει τις ρίζες της σε αυτή την απόδειξη της νευρωνικής ασυνέχειας από τον Ramon y Cajal. Αργότερα, από τον Pavlov, αλλά ιδιαίτερα από τους Hebb, Sherrington και Kandel, εξετάζεται η ιδέα της αυξομείωσης της συναπτικής αποτελεσματικότητας ως υπόβαθρο της μάθησης και μνήμης. Αυτού του είδους η πλαστικότητα παρατηρήθηκε για πρώτη φορά στα κύτταρα του ιππόκαμπου αρχικά ως Μακρόχρονη Ενδυνάμωση-LTP (Bliss & Lψmo, 1973), αργότερα ως Μακρόχρονη Αποδυνάμωση-LTD (Levy & Stewart, 1979) και τέλος ως δυναμική κατάσταση αντιστρέψιμης συναπτικής πλαστικότητας (Dudek & Bear,1992; Kirkwood & Bear,1994; Mayford et al,1995).
Οι μηχανισμοί που επιτρέπουν μια τέτοια πλαστική συναπτική σύνδεση εξακολουθούν να αποτελούν το αντικείμενο έντονων συζητήσεων. Αρχικό σημείο διαφωνίας ήταν το άν ο εστιακός τόπος της πλαστικότητας ήταν προ- ή μετα-συναπτικός. Πολύ πιο πολύπλοκο να απαντηθεί είναι το ερώτημα του συνολικού μηχανισμού αυτής της πλαστικότητας. Κατά καιρούς έχουν προταθεί ανατομικά, μοριακά, δομικά και ηλεκτρικά μοντέλα που όμως αδυνατούν να διατηρήσουν την περιγραφική τους ικανότητα για όλα τα φαινόμενα που συνοδεύουν τη συναπτική πλαστικότητα. Η παρούσα εργασία μελετά τα μέχρι σήμερα πειραματικά δεδομένα και παρατηρήσεις και προσπαθεί όχι μόνο να περιγράψει το μηχανισμό που επιφέρει την Μακρόχρονη Αποδυνάμωση αλλά και να της αποδώσει σαφή λειτουργικό ρόλο.
Α. Ηλεκτροφυσιολογία
1. Εγκεφαλικές τομές
Επίμυες τύπου Wistar (n=11, 4αρσ. και 7θηλ.) αναπτύχθηκαν σε καλά φωτιζόμενο περιβάλλον με κανονικές τεχνητές εναλλαγές ημέρας/νύχτας (12/12h αντίστοιχα). Τα ζώα θυσιάστηκαν ως νεαροί ενήλικες (ηλικίας 4 μηνών), ένα κάθε φορά, αρχικά με βαθιά αναισθησία με αιθέρα και αποκεφαλισμό τους με γκιλοτίνα, σε διαφορετικό χώρο από αυτόν της φύλαξης των ζώων και του εργαστηρίου ώστε να μην επέλθει stress τρόμου στα υπόλοιπα ζώα. Το δέρμα της κεφαλής των ζώων κόπηκε ινιορινικά και το κρανίο τους διανοίχτηκε κατά μήκος της βρεγματικής ραφής με προσοχή να μην τραυματιστεί ο υποκείμενος εγκεφαλικός ιστός. Οι πλήρεις εγκέφαλοί τους αφαιρέθηκαν μέσα σε 90sec και βυθίστηκαν αμέσως σε πολύ ψυχρό (2-4 °C) τεχνητό εγκεφαλονωτιαίο υγρό (artificial cerebrospinal fluid ή ΑCSF): 124mΜ NaCl, 4mM KCl, 2mM MgSO4, 2mM CaCl2, 1.25mM NaH2PO4, 26mM NaHCO3 και 10mM Glucose, συνεχώς υπεροξυγονωμένο (95% O2 και 5% CO2) με αναβλύζον οξυγόνο και σε pH που ποτέ δεν ξεπέρασε το 7.34 (+/- 0.02), σύμφωνα με την ευρύτερα αποδεκτή σύστασή του (Brown et al, 1988; Bliss & Collingridge,1993).
Μετά την αποκοπή των οσφρητικών λοβών και την στεφανιαία αφαίρεση των ακραίως μετωπιαίων και παρεγκεφαλιδικών περιοχών, οι κυρίως εγκέφαλοι διαχωρίστηκαν στη μέση επιμήκη σχισμή και στο μεσολόβιο, στη συνέχεια το κάθε ημισφαίριο σταθεροποιήθηκε με στιγμιαία εποξική κόλλα (Logo) στη βάση κοπής ενός οριζόντιου μικροτόμου δόνησης (vibratome) McIWAAN R500 και ελήφθησαν εγκάρσιες τομές του ιππόκαμπου πάχους 400-450μm. Οι τομές αυτές μεταφέρθηκαν αμέσως σε διηθητικό χαρτί που βρισκόταν σε θάλαμο ύγρανσης με αλληλεπίδραση (humidifying interface-type chamber) κατασκευασμένο από Plexiglas όπου, πριν οποιοδήποτε πειραματικό σχήμα, διατηρήθηκαν επί 1.5 ώρες διαποτιζόμενες από ACSF διηθητικής ροής στους 37 °C (+/- 0.2 °C) προκειμένου η ηλεκτρική δραστηριότητα των νευρώνων να επανέλθει στα φυσιολογικά επίπεδα (Brown et al.1988; Chen et al.1999).
2. Ηλεκτρική διέγερση
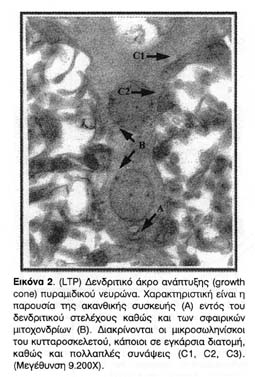
Σε όλα τα ηλεκτροφυσιολογικά πειράματα που πραγματοποιήθηκαν στη συνέχεια, ως ηλεκτρόδιο διέγερσης χρησιμοποιήθηκε ένα διπλό, ελικοειδώς περιπλεγμένο και μονωμένο ατσάλινο καλώδιο (διαμέτρου 50μm) που τοποθετήθηκε στην περιοχή των παράπλευρων κλωνίων του Schaffer (Schaffer collaterals) για την διπολική διέγερση των δενδριτικών συνάψεων της ακτινωτής στιβάδας (stratum radiatum) της CA1 περιοχής του ιππόκαμπου (Εικόνα 2). Ως ηλεκτρόδια καταγραφής τραβήχτηκαν τριχοειδή σωληνοειδή από βορικοπυριτικό ύαλο (borosilicate glass) σε Flamming-Brown P-80 micropipette puller, Sutter Instruments. Η αντίσταση των ηλεκτροδίων αυτών, μετά την πλήρωσή τους με 2Μ οξικό κάλιο, ήταν μεταξύ 80 και 120ΜΩ για τα ηλεκτρόδια ενδοκυττάριας καταγραφής και μεταξύ 1 και 10ΜΩ για τα ηλεκτρόδια εξωκυττάριας καταγραφής. Ηλεκτρόδια μικρότερης αντίστασης απορρίπτονταν.
Σε όλη τη διάρκεια των πειραμάτων οι τομές διατηρήθηκαν διαποτιζόμενες από ACSF το οποίο περνούσε από ψηφιακό υδατόλουτρο ώστε να παραμένει στους 37 °C (+/-0.2 °C), για μέγιστο 18 ωρών μετά την απαρχή των ηλεκτροφυσιολογικών πειραμάτων.
Η τοποθέτηση των ηλεκτροδίων γινόταν με τη βοήθεια μηχανικών μικροχειριστών (micromanipulators) ZEISS AV-921 κάτω από μεγέθυνση (x10) οπτικού ανατομικού μικροσκοπίου NIKON SM2-10A και ψυχρό φωτισμό αλογόνου. Όλη η πειραματική διάταξη είχε τοποθετηθεί πάνω σε χαλύβδινη τράπεζα με ελαστικό μηχανισμό μηδενικής αδράνειας και μέσα σε έναν κλωβό Faraday ηλεκτρικής απομόνωσης ο οποίος κάλυπτε όλο το χώρο της ηλεκτροφυσιολογικής κυτταρικής μελέτης.
Το ηλεκτρικό σήμα, που θα έφτανε στον ιστό μέσω του ηλεκτροδίου διέγερσης, προγραμματιζόταν σε ηλεκτρονικό διεγέρτη τύπου Master-8, ο οποίος δεχόταν εξωτερικά τη πρώτη πυροδότηση (triggering signal). Για την πρόκληση σταθερής νευρωνικής βασικής απόκρισης, η οποία στη συνέχεια έπαιζε το ρόλο του μάρτυρα προκειμένου να αποδειχτεί η μεταβολή της συναπτικής πλαστικότητας, ο διεγέρτης έδινε τετράγωνο παλμό διάρκειας 10msec και συχνότητας 10Ηz επί τουλάχιστον 30min. Για την φάση της πρόκλησης LTP ο διεγέρτης προγραμματίστηκε να δίνει ερέθισμα υψηλόσυχνης ριπής (HFS) τύπου θ (θ-burst stimulation protocol), δηλαδή μία ακολουθία ριπών 10 τετράγωνων παλμών διάρκειας 10msec και συχνότητας 100Hz, με κάθε ριπή παλμών να διαχωρίζεται με 200msec από την επόμενη, για σύνολο 900 παλμών. Για την φάση της πρόκλησης LTD ο διεγέρτης προγραμματίστηκε να δίνει ερέθισμα χαμηλόσυχνης ριπής (LFS) δηλαδή μία ακολουθία τετράγωνων παλμών διάρκειας 10msec και συχνότητας 1Ηz, με κάθε ριπή παλμών να διαχωρίζεται με 200msec από την επόμενη, για σύνολο 900 παλμών (Staubli & Zi, 1996; Kirkwood et al, 1996; Petersen et al,1998).
3. Ηλεκτρονική καταγραφή και ανάλυση
Η λήψη και καταγραφή της ηλεκτρικής απόκρισης του ιστού έγινε μέσω ενός Axolab1100 Interface που μετέτρεπε το σήμα από τον προενισχυτή AxoClamp-2A σε ψηφιακό, προκειμένου να αναγνωριστεί και εν συνεχεία να αναλυθεί από το λογισμικό Pclamp 5.03 (AXON Instuments) που έτρεχε σε προσωπικό υπολογιστή με επεξεργαστή Intel Pentium II των 550Hz. Ταυτόχρονα με την επεξεργασία και καταγραφή της ηλεκτρικής απόκρισης του ιστού η μορφή του απεικονιζόταν σε παλμογράφο ώστε να γίνουν αμέσως αντιληπτά τυχόν πειραματικά προβλήματα, όπως για παράδειγμα τυχόν θραύση του ηλεκτροδίου καταγραφής.
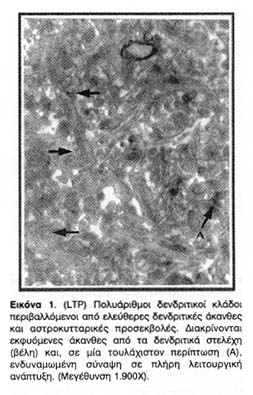
Το ηλεκτρόδιο διέγερσης τοποθετούνταν στην περιοχή των παράπλευρων κλωνίων του Schaffer (Schaffer collaterals) που περιέχει άξονες των πυραμιδικών νευρώνων της CA3 περιοχής του ιππόκαμπου. Τα ηλεκτρόδια καταγραφής, εξωκυττάρια και ενδοκυττάρια, τοποθετούνταν στην περιοχή της ακτινωτής στιβάδας (stratum radiatum) η οποία περιέχει πολλούς και πλούσια διακλαδωμένους δενδρίτες πυραμιδικών κυττάρων της CA1 περιοχής (Εικόνα 1).
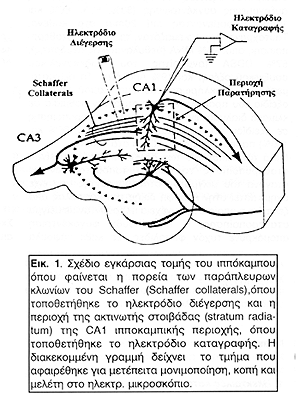
Οι ενδοκυττάριες καταγραφές μετρούν τη διαφορά δυναμικού της εσωτερικής πλευράς της νευρωνικής μεμβράνης σε σχέση με το νευροπιληματικό χώρο. Καθώς αυτές δεν προσφέρονται για απόδειξη της πρόκλησης LTP ή LTD σε έναν πληθυσμό νευρώνων χρησιμοποιήθηκαν μόνο ως επιβεβαίωση της κυτταρικής απόκρισης και μεταβολής της συναπτικής πλαστικότητας ανάλογα με τα χαρακτηριστικά του προσυναπτικού ερεθίσματος. Οι εξωκυττάριες καταγραφές όμως μετρούν τη συνολική απόκριση ενός πληθυσμού νευρώνων και αυτές χρησιμοποιήθηκαν για την πειραματική απόδειξη της αύξησης ή μείωσης της συναπτικής πλαστικότητας.
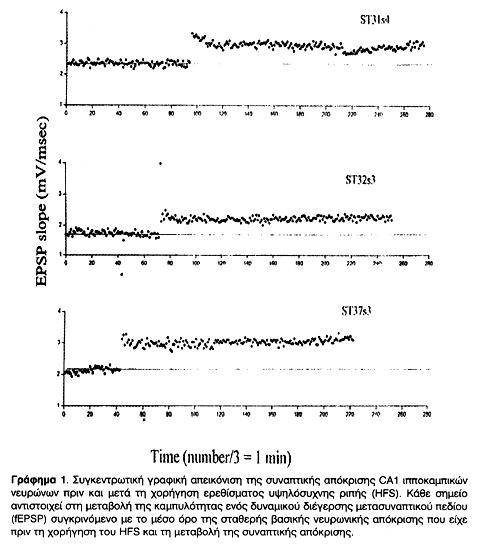
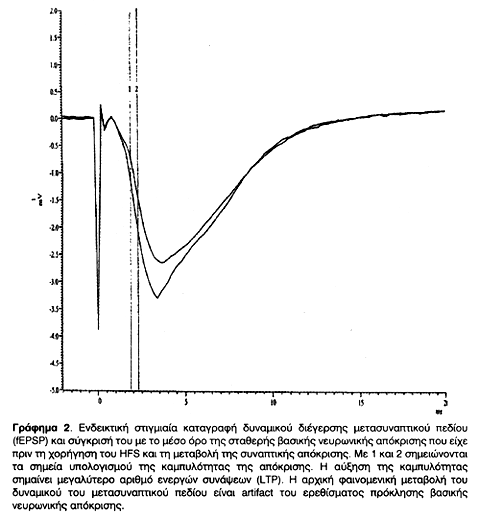
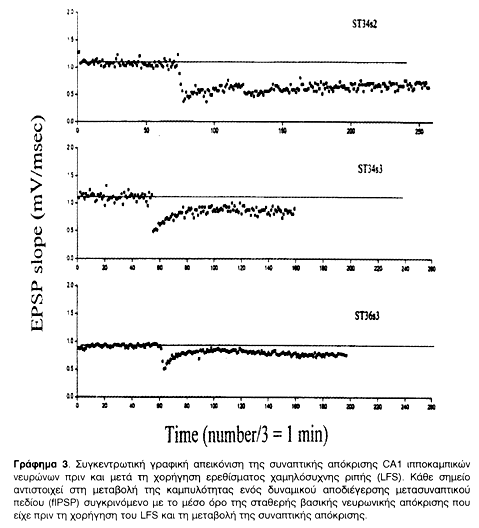
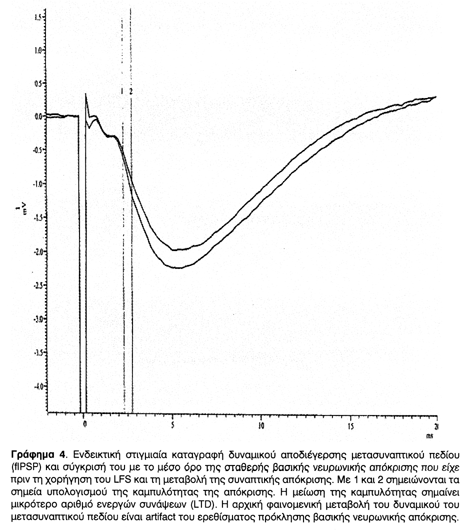
Μετά την ολοκλήρωση της χορήγησης HFS ή LFS καταγραφόταν το δυναμικό διέγερσης ή αποδιέγερσης μετασυναπτικού πεδίου (fEPSP ή fIPSP) που προκαλείται από ερέθισμα όμοιο με εκείνο που είχε χορηγηθεί για την πρόκληση σταθερής βασικής νευρωνικής απόκρισης (μάρτυρας). Η σύγκριση της καμπυλότητας των δυναμικών πριν και μετά τη χορήγηση του ερεθίσματος δίνει τη μεταβολή της συναπτικής απόκρισης. Η αυξημένη καμπυλότητα σημαίνει μεγαλύτερο αριθμό ενεργών συνάψεων, και άρα μείωση του κατωφλίου εναλλαγής θm, ενώ η μειωμένη σημαίνει μικρότερο αριθμό λόγω της ανύψωσης του θm. Αυτές οι μεταβολές στην πλαστικότητα απεικονίστηκαν σε Γραφήματα, στιγμιαία (Γραφήματα 2, 4) και συγκεντρωτικά (Γραφήματα 1 & 3). Οι καταγραφές συνεχίστηκαν μέχρι και 18 ώρες μετά τη χορήγηση του HFS ή LFS και διαπιστώθηκε η σταθερότητα των συναπτικών μεταβολών πλαστικότητας.
Με το παραπάνω πειραματικό σχήμα έγινε δυνατό να επιβεβαιωθεί η πρόκληση και σταθεροποίηση LTP και LTD σε δενδριτικές συνάψεις της CA1 περιοχής του ιππόκαμπου, τομών οι οποίες στη συνέχεια μελετήθηκαν στο ηλεκτρονικό μικροσκόπιο.
Β. Ηλεκτρονική μικροσκοπία
1. Προετοιμασία, κοπή και βαφή δειγμάτων
Αμέσως μετά την ολοκλήρωση των ηλεκτρο-φυσιολογικών πειραμάτων, από τις ιπποκαμπικές τομές αφαιρέθηκαν ιστοτεμάχια που περιέκλειαν την περιοχή όπου έγινε η καταγραφή της συναπτικής μεταβολής. Τα ιστοτεμάχια αυτά στη συνέχεια μεταφέρονταν και παρέμεναν για τουλάχιστον 48 ώρες σε μονιμοποιητικό διάλυμα Sotelo: 25% γλουταραλδεϋδη, 6% παραφορμαλδεϋδη, 1% CaCl και ίχνη NaOH σε ρυθμιστικό διάλυμα κακοδυλικού οξέος (cacodylate buffer) (pH 7.34, +/- 0,02).
Μετά τη παρέλευση 48 ωρών τα ιστοτεμάχια ξεπλένονταν με άφθονο κακοδυλικό buffer, κατατέμνονταν σε μικρότερα τεμάχια 2x2mm (Εικόνα 2) και στη συνέχεια μεταφέρονταν σε διάλυμα τετροξειδίου του οσμίου (1%) σε κακοδυλικό buffer όπου παρέμειναν για 30min. Τα τεμάχια αυτά στη συνέχεια ξεπλένονταν με άφθονο κακοδυλικό buffer μέχρι να σταματήσει να απελευθερώνεται μπλε χρωστική από το όσμιο που βρέθηκε σε περίσσεια. Μετά την πλύση τα δείγματα μεταφέρονταν σε υδάτινο διάλυμα ακετόνης (70%) όπου σε κάποιες περιπτώσεις παρέμεναν μέχρι και 24 ώρες.
Την επόμενη ημέρα τα δείγματα αφυδατώνονταν σε κλίμακα υδατικών διαλυμάτων ακετόνης αυξανόμενης συγκέντρωσης (70%, 80%, 90% και 100%) παραμένοντας επί 10min στο καθένα. Στη συνέχεια διαποτίζονταν βαθμιαία σε κλίμακα διαλυμάτων οξειδίου του προπυλενίου σε ακετόνη αυξανόμενης συγκέντρωσης (33%, 50%, 66% και 100%) παραμένοντας επί 20min στο καθένα. Ταυτόχρονα παρασκευαζόταν το μείγμα σταθεροποίησης από: 24% MNA (μεθυλαναδικός ανυδρίτης), 57% DDSA (ανυδρίτης του δωδεκανυλικού σουξινικού οξέος) και 19% Αραλδίτη. Τα δείγματα, μετά την ολοκλήρωση του εμποτισμού τους με οξείδιο του προπυλενίου, εμποτίζονταν βαθμιαία σε κλίμακα διαλυμάτων του μείγματος σταθεροποίησης σε οξειδίου του προπυλενίου (33%, 50%, 66% και 100%) παραμένοντας επί 30min στο καθένα.
Μετά τον εμποτισμό των δειγμάτων από το μείγμα σταθεροποίησης το καθένα τοποθετούνταν σε σταγόνα του μείγματος σταθεροποίησης που είχε σχηματιστεί στην κορυφή δικής του κάψουλας από ζελατίνη η οποία στη συνέχεια πληρούνταν με μείγμα σταθεροποίησης. Η επακόλουθη φυγοκέντριση απομάκρυνε τυχόν φυσαλίδες. Σε κάθε κάψουλα προσθέτονταν και σταγόνες DMP-30 (2,4,6-τρισφενολικο-διμεθυλοαμινομεθύλιο) (16 σταγόνες ανά 28ml μείγματος) το οποίο καταλύει την ταχύτερη πήξη του μείγματος. Οι κάψουλες στη συνέχεια μεταφέρονταν σε κλίβανο 60oC όπου παρέμεναν επί 48 ώρες μέχρι να στερεοποιηθεί το μείγμα σταθεροποίησης. Μετά τη στερεοποίηση οι κάψουλες και τα δείγματα, είναι έτοιμα για τομή και χρώση.
Για την κοπή των δειγμάτων χρησιμοποιήθηκε ο ηλεκτρονικός υπερμικροτόμος REICHERT JUNG Ultracute. Από κάθε δείγμα κόπηκαν αρχικά ημίλεπτες τομές (πάχους 0.5-1.0 μm) οι οποίες βάφηκαν με διάλυμα κυανού της τολουϊδίνης (1%) και μελετήθηκαν σε οπτικό μικροσκόπιο, ZEISS Universal, ώστε να εντοπιστεί το καλύτερο πεδίο για υπερμικροσκοπική μελέτη. Στη συνέχεια από κάθε δείγμα κόπηκαν υπέρλεπτες τομές (πάχους 15-30nm) οι οποίες τοποθετήθηκαν πάνω σε χάλκινα πλέγματα παρατήρησης.
Η τελική χρώση των υπέρλεπτων τομών έγινε με την τοποθέτησή τους στην επιφάνεια σταγόνων υδατικού και αλκοολικού (50%) διαλύματος οξικού ουρανυλίου (3%) επί 15min, πλύση με αιθυλική αλκοόλη (50%) και στη συνέχεια με απεσταγμένο νερό, τοποθέτησή τους στην επιφάνεια σταγόνων διαλύματος κιτρικού μολύβδου (1%) επί 3min και, τέλος, στέγνωμα με εξάτμιση πάνω σε διηθητικό χαρτί.
2. Παρατήρηση και φωτογράφηση
Όλες οι παρατηρήσεις και φωτογραφήσεις υπερμικροσκόπησης έγιναν σε ηλεκτρονικό μικροσκόπιο ZEISS EM 95-2. Τα φιλμ που χρησιμοποιήθηκαν ήταν ασπρόμαυρα μικροκοκκικά φιλμ της AGFA τα οποία εμφανίστηκαν με developer AGFA, Rodinal special, σταθεροποιήθηκαν με AGFA Acidofix και εκτυπώθηκαν σε πλαστικό φωτογραφικό χαρτί KODAK. Οι φωτογραφίες που μελετήθηκαν φαίνεται να επιβεβαιώνουν το προτεινόμενο συνδυαστικό μοντέλο (Φωτογραφίες 1-4).
 |
|
 |
|
 |
 |
|
 |
|
 |
Παρατηρήσεις
1. Ηλεκτρικές καταγραφές
Σκοπός των ηλεκτρικών καταγραφών ήταν η απόδειξη της μεταβολής της συναπτικής πλαστικότητας και η πρόκληση σταθερής LTP ή LTD στους ιπποκαμπικούς ιστούς που στη συνέχεια θα μελετούνταν κάτω από το ηλεκτρονικό μικροσκόπιο. Τόσο η συναπτική ενδυνάμωση όσο και η αποδυνάμωση ολοκληρώθηκαν με επιτυχία και παρέμειναν σταθερές για αρκετές ώρες, όπως φαίνεται και από τα συγκεντρωτικά Γραφήματα 1 και 3.
Για την πρόκληση σταθερής νευρωνικής βασικής απόκρισης, η οποία στη συνέχεια έπαιζε το ρόλο του μάρτυρα προκειμένου να αποδειχτεί η μεταβολή της συναπτικής πλαστικότητας, ο διεγέρτης έδινε βασικό ερέθισμα ελέγχου. Για την φάση της πρόκλησης LTP ο διεγέρτης προγραμματίστηκε να δίνει ερέθισμα υψηλόσυχνης ριπής (HFS) στα 100Hz ενώ για την φάση της πρόκλησης LTD ο διεγέρτης να δίνει ερέθισμα χαμηλόσυχνης ριπής (LFS) στα 1Ηz.
Μετά την ολοκλήρωση της χορήγησης HFS ή LFS καταγραφόταν το δυναμικό διέγερσης ή αποδιέγερσης μετασυναπτικού πεδίου (fEPSP ή fIPSP) που προκαλείται από ερέθισμα όμοιο με εκείνο που είχε χορηγηθεί για την πρόκληση σταθερής νευρωνικής βασικής απόκρισης (μάρτυρας). Η σύγκριση της καμπυλότητας των δυναμικών πριν και μετά τη χορήγηση του ερεθίσματος δίνει τη μεταβολή της συναπτικής απόκρισης, όπως φαίνεται στα Στιγμιαία Γραφήματα (2, και 4). Η αυξημένη καμπυλότητα σημαίνει μεγαλύτερο αριθμό ενεργών συνάψεων, και άρα μείωση του θm, ενώ η μειωμένη σημαίνει μικρότερο αριθμό λόγω της ανύψωσης του θm. Αυτές οι μεταβολές στην πλαστικότητα απεικονίστηκαν σε Συγκεντρωτικά Γραφήματα (1 & 3). Οι καταγραφές συνεχίστηκαν μέχρι και 18 ώρες μετά τη χορήγηση του HFS ή LFS και διαπιστώθηκε η σταθερότητα των συναπτικών μεταβολών πλαστικότητας. Με τον τρόπο αυτό έγινε δυνατό να επιβεβαιωθεί η πρόκληση και σταθεροποίηση LTP και LTD σε δενδριτικές συνάψεις της CA1 περιοχής του ιππόκαμπου τομών οι οποίες στη συνέχεια μελετήθηκαν στο ηλεκτρονικό μικροσκόπιο.
2. Φωτογραφήσεις ηλεκτρονικής μικροσκόπησης
Από τις φωτογραφίες ηλεκτρονικής μικροσκόπησης που ελήφθησαν παρατίθενται οι πλέον χαρακτηριστικές. Όλες προέρχονται από τη CA1 ιπποκαμπική περιοχή και συγκεκριμένα από την περιοχή που περιέβαλλε το σημείο της τοποθέτησης του ηλεκτροδίου καταγραφής δηλαδή μία περιοχή της ακτινωτής στιβάδας (stratum radiatum), η οποία περιέχει πολλούς και πλούσια διακλαδωμένους δενδρίτες πυραμιδικών κυττάρων της CA1 περιοχής (Εικόνα 1).
Οι φωτογραφίες χωρίστηκαν σε δύο ομάδες, στην πρώτη παρουσιάζονται οι νευρώνες ιπποκαμπικών τομών που είχαν δεχτεί ερέθισμα υψηλόσυχνης ριπής και είχαν ενδυναμώσει τη συναπτική τους απόκριση (LTP) (Φωτογραφίες 1-3) και στη δεύτερη οι νευρώνες ιπποκαμπικών τομών που είχαν δεχτεί ερέθισμα χαμηλόσυχνης ριπής και είχαν αποκτήσει αποδυναμωμένη απόκριση (LTD) (Φωτογραφίες 4-6). Μέσα σε κάθε ομάδα οι φωτογραφίες έχουν διαταχθεί συνήθως σε σειρά αυξανόμενης μεγέθυνσης ώστε να γίνει ευκολότερη και σαφέστερη η παρουσίαση των δομικών μεταβολών που παρατηρήθηκαν.
Τέλος, χρησιμοποιήθηκαν βέλη με γραμματοσήμανση ως βοηθήματα παρουσίασης των παρατηρήσεων που αναγράφονται στα υπομνήματα των φωτογραφιών για ακριβέστερη περιγραφή. Τα συμπεράσματα των παρατηρήσεων συνοψίστηκαν μετά την παρουσίαση των φωτογραφικών δεδομένων και η σημασία τους αναλύεται στη συζήτηση όπου παρατίθενται και τα σχετικά συμπεράσματα. Η εξαγωγή συμπερασμάτων από τη μελέτη φωτογραφιών ηλεκτρονικής μικροσκοπίας συνοδεύεται πάντα από τον κίνδυνο, μοναδικές παρατηρήσεις να αναχθούν σε γενικευμένα συμπεράσματα. Όταν όμως το ίδιο φαινόμενο καθίσταται επανειλημμένα αντιληπτό σε μεγάλο αριθμό δειγμάτων, τότε οι παρατηρήσεις αντικατοπτρίζουν ένα πραγματικό φαινόμενο και η μελέτη κάτω από το ηλεκτρονικό μικροσκόπιο επιτρέπει τον εντοπισμό λεπτομερειών που θα διέφευγαν με μελέτη διαφορετικής προσέγγισης.
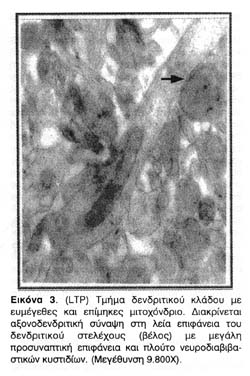
Κοινό χαρακτηριστικό των συνάψεων των ενδυναμωμένων νευρώνων είναι η αύξηση όχι μόνο της συνολικής συναπτικής επιφάνειας αλλά και της λειτουργικής μετασυναπτικής επιφάνειας της μετασυναπτικής πύκνωσης (PSD), η οποία φαίνεται ως ηλεκτρονιακά πυκνή (σκουρόχρωμη) πάχυνση της μετασυναπτικής μεμβράνης και η οποία αντιστοιχεί σε μεμβρανική περιοχή πλούσια σε υποδοχείς και διαύλους οργανωμένους σε κβαντικές δέσμες.
Όπως φαίνεται από τις φωτογραφίες 1, 2 και 3 η πρόκληση LTP οδηγεί τις άκανθες, των οποίων οι συνάψεις ενδυναμώνονται, να διογκωθούν, και να αυξήσουν έτσι την αποτελεσματικότητα των συνάψεων τους, με την προέκταση της λειτουργικής επιφάνειάς τους. Επιπλέον, οι φωτογραφίες αυτές είναι ενδεικτικές της δενδριτικής ανάπτυξης νέων κλάδων και ακανθικών προεκβολών που προκαλείται από την ενδυνάμωση των ιπποκαμπικών συνάψεων. Αυτές οι παρατηρήσεις, βέβαια, απλώς επιβεβαίωσαν παλαιότερα συμπεράσματα για τις δομικές και ανατομικές μετατροπές που επιφέρει η πρόκληση LTP στους δενδρίτες της CA1 περιοχής του ιππόκαμπου και ήταν ήδη γνωστές από παλαιότερες εργασίες.
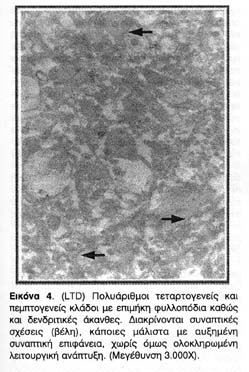
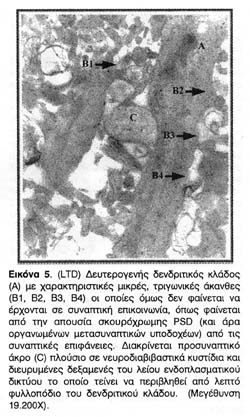
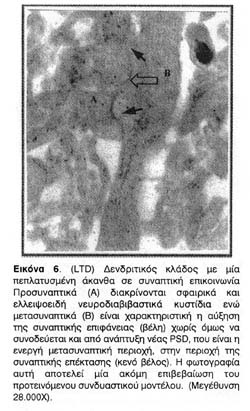
Ιδιαίτερο ενδιαφέρον παρουσίασαν οι δομικές μετατροπές που παρατηρήθηκαν στους δενδρίτες ιπποκαμπικών τομών που είχαν υποστεί συναπτική αποδυνάμωση. Ενώ οι δομικές μετατροπές στην περίπτωση της LTP έχουν θεμελιωθεί και επανειλημμένα αποδειχτεί, μέχρι σήμερα δεν είχαν αναφερθεί δομικές μετατροπές στην περίπτωση της LTD. Οι φωτογραφίες 4, 5, και ιδίως η 6 απεικονίζουν σαφέστατα την επέκταση της μετασυναπτικής επιφάνειας σε αποδυναμωμένες συνάψεις αλλά όχι και της PSD, δηλαδή της ενεργού λειτουργικής μετασυναπτικής περιοχής τους.
Επιπλέον, όπως φαίνεται από τη δεύτερη ομάδα φωτογραφιών, παρ' όλο που ο νευροπηλιματικός χώρος βρίθει με δενδριτικούς κλώνους και αξονικά προσυναπτικά άκρα, η αποδυνάμωση φαίνεται ότι επιφέρει έναν δισταγμό στον σχηματισμό συναπτικής επικοινωνίας με αποτέλεσμα τη μείωση των συναπτικών σχέσεων.
Τα συμπεράσματα αυτών των παρατηρήσεων, σε συνδυασμό με την μετα-αναλυτική μελέτη των συσσωρευμένων πειραματικών αποτελεσμάτων των 30 τελευταίων χρόνων, επέτρεψε τη διαμόρφωση ενός ηλεκτροδομικού μοντέλου για ην μακρόχρονη αποδυνάμωση.
Συζήτηση
Η μελέτη της συναπτικής πλαστικότητας ως το κυτταρικό υπόστρωμα της μνήμης, μιας αναδυόμενης ιδιότητας του εγκεφάλου, δεν είναι εύκολο να δώσει σαφείς απαντήσεις αν επικεντρωθεί στα επιμέρους χαρακτηριστικά της, κυρίως γιατί εμφανίζεται με πολλές μορφές, τόσο στις διάφορες περιοχές του εγκεφάλου όσο και ανάμεσα στα διαφορετικά πεδία του ίδιου του ιππόκαμπου. Έχουν περάσει 30 χρόνια από την ανακάλυψη της πρώτης μορφής της, ως LTP το 1973, και από τότε έχει συσσωρευθεί ένας τεράστιος όγκος πειραματικών δεδομένων, κάποια συμπληρωματικά ενώ κάποια άλλα αντικρουόμενα. Ένα από τα ερωτήματα που επέμεναν να παραμένουν αναπάντητα ήταν ο μηχανισμός και ο λειτουργικός ρόλος της LTD.
Η προσέγγιση της συναπτικής πλαστικότητας με την παρούσα εργασία βασίζεται κυρίως σε δεδομένα από την CA1 περιοχή του ιππόκαμπου αλλά προσεγγίζει το φαινόμενο συνολικά, λαμβάνοντας υπόψη και τα δεδομένα άλλων περιοχών του εγκεφάλου. Στόχος ήταν να εντοπιστούν οι κοινοί μηχανισμοί, οι οποίοι πιθανόν να εκφράζονται μέσω διαφορετικών μορίων ή κυτταρικών σχηματισμών κατά περίπτωση.
Κεντρικής σημασίας φαίνεται να είναι οι αυξομειώσεις των συγκεντρώσεων των ιόντων Ca++ σε διάφορες μικροπεριοχές του νευρωνικού κυττάρου. Σε αυτές μπορεί να αναζητηθεί η ερμηνεία πολλών φαινομένων όπως η πρόκληση και έκφραση της LTD.
Η προσαγωγή ερεθίσματος υψηλόσυχνης ριπής (ΗFS) in vitro, αλλά και η μαζική συναπτική συνενεργοποίηση πολλών συνάψεων του ίδιου δενδριτικού κλάδου ενός νευρώνα in vivo, ανυψώνει αρκετά το δυναμικό της μεμβράνης (Vm) των δενδριτικών ακάνθων με αποτέλεσμα την απομάκρυνση των ιόντων Mg++ από το δίαυλο των υποδοχέων NMDA, την ενεργοποίησή τους και την εισροή ιόντων Ca++. Στην εισροή ιόντων Ca++ στην περισυναπτική περιοχή μέσω των υποδοχέων NMDA προστίθενται και τα ιόντα Ca++ που απελευθερώνονται από τις δεξαμενές του λείου ενδοπλασματικού δικτύου μέσω της δράσης της φωσφορικής ινοσιτόλης-3 (ΙΡ3), η οποία αποτελεί προϊόν της διάσπασης φωσφολιπιδίων κάτω από την επίδραση της φωσφολιπάσης, που είχε ενεργοποιηθεί με τη σειρά της από G-πρωτεΐνες. Αυτή η αυξημένη μικροσυγκέντρωση ιόντων Ca++ ενεργοποιεί την CaM-KII η οποία, αφενός, πυροδοτεί το σχηματισμό και την απελευθέρωση ανάδρομων αγγελιοφόρων, όπως το μονοξείδιο του αζώτου και το αραχιδονικό οξύ, αφετέρου αυξάνει την αποτελεσματικότητα των υποδοχέων AMPA ως διαύλων ιόντων Ca++, οι οποίοι έτσι παρατείνουν την αυξημένη εισροή ιόντων Ca++ στη συγκεκριμένη περιοχή. Τα παραπάνω γεγονότα λειτουργούν συνεργικά, καθώς ένας από τους ρόλους των ανάδρομων αγγελιοφόρων είναι η αύξηση της ποσότητας και παράτασης του χρόνου απελευθέρωσης του νευροδιαβιβαστή από το προσυναπτικό άκρο. Παράλληλα έχει ξεκινήσει και ο πολυμερισμός των ινιδίων ακτίνης, τόσο από την αυξημένη μικροσυγκέντρωση Ca++ όσο και από τη δράση της CaM-KII και της PKC σε πρωτεΐνες, όπως η καλδεσμόνη που απομακρύνονται από τα θετικά άκρα των ινιδίων. Ο πολυμερισμός αυτός ωθεί τη μεμβράνη της μετασυναπτικής επιφάνειας σε επιμήκυνση, δρομολογώντας έτσι τις γνωστές δομικές μετατροπές της μετασυναπτικής επιφάνειας. Εδώ θα πρέπει να σημειωθεί πως ένας δεύτερος ρόλος που έχει αποδοθεί στους ανάδρομους αγγελιοφόρους, είναι η πρόκληση προσυναπτικών δομικών μεταβολών έτσι ώστε να εξακολουθεί να υφίσταται λειτουργική σύζευξη μεταξύ των δύο συναπτικών επιφανειών.
Αν το προσυναπτικό ερέθισμα ήταν αρκετά μεγάλης συχνότητας και διάρκειας ή προέκυψε από ταυτόχρονη πολλαπλή συνενεργοποίηση πολλών συνάψεων του ίδιου δενδριτικού κλάδου, η μεταβολή του δυναμικού της μεμβράνης (Vm) θα επεκταθεί ως τη βάση της άκανθας και η εισροή των ιόντων Ca++ θα παραταθεί με την ενεργοποίηση των VDCC που έχουν εντοπισθεί εκεί. Στο δενδριτικό στέλεχος υπάρχουν πολυριβοσώματα και mRNA για τη σύνθεση υποδοχέων, ιοντικών διαύλων και μεμβρανικών πρωτεϊνών. Η διαρροή ιόντων Ca++ σε αυτό επιτάσσει τη μετάφραση των mRNA και την απορρόφηση των πρωτεϊνικών προϊόντων στην άκανθα υπό ενδυνάμωση. Τα προϊόντα αυτά θα καταστήσουν λειτουργικά ενεργή την αυξημένης επιφάνειας σύναψη, κάτι που συνεπάγεται συναπτική ενδυνάμωση καθώς αυξάνεται η επιφάνεια διαυλικής αποκρισιμότητας και έτσι ένα προσυναπτικό ερέθισμα της ίδιας συχνότητας και έντασης δημιουργεί ευκολότερα μεγαλύτερη απόκριση. Τα μόρια αυτά εισέρχονται στη μεμβράνη με τη σύντηξη των κυστιδίων μεταφοράς της δεξαμενής υποδοχέων (ΔΙΑΓΡΑΜΜΑ 1).

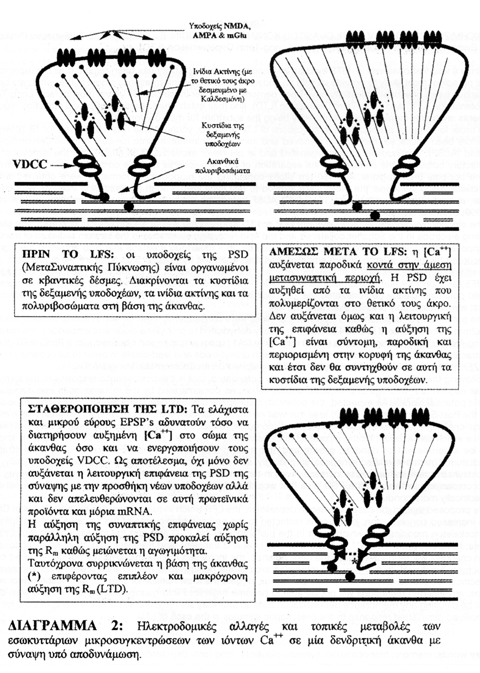
Στην περίπτωση προσαγωγής ερεθίσματος χαμηλόσυχνης ριπής (LFS) in vitro, αλλά και της μερικής συναπτικής συνενεργοποίησης κάποιων μόνο συνάψεων του ίδιου δενδριτικού κλάδου ενός νευρώνα in vivo, η αύξηση της μικροσυγκέντρωσης των ιόντων Ca++ στην περισυναπτική περιοχή, λόγω της τοπικής απόφραξης των υποδοχέων NMDA, είναι αρκετή για να ξεκινήσει ο πολυμερισμός των ινιδίων ακτίνης. Ο πολυμερισμός αυτός αυξάνει τη μεμβράνη της συναπτικής επιφάνειας αλλά δεν καθίσταται λειτουργικά ενεργός, καθώς δεν συνοδεύεται από απορρόφηση νέων υποδοχέων και ιοντικών διαύλων από το δενδριτικό στέλεχος, κάτι που απαιτεί παρατεταμένη εισροή ιόντων Ca++ και διάχυσή τους στη βάση της άκανθας. Με αυτό το τρόπο, η άκανθα χάνει το λειτουργικό της σχήμα και μετέπειτα προσυναπτικά ερεθίσματα θα τύχουν αποδυναμωμένης συναπτικής απόκρισης καθώς η αύξηση της επιφάνειας χωρίς αντίστοιχη αύξηση της αγωγιμότητας συνεπάγεται αύξηση της ηλεκτρικής αντίστασης της μεμβράνης (Rm).
Η έκταση των δομικών μετατροπών θα καθορίσει τη διάρκεια της αποδυναμωμένης συναπτικής απόκρισης και ανάλογα θα έχει ως αποτέλεσμα μόνο βραχύχρονη (STD) ή μακρόχρονη (LTD) αποδυνάμωση, στην περίπτωση που συμβεί περίσφιξη της βάσης της άκανθας αυξάνοντας ακόμα περισσότερο την Rm (ΔΙΑΓΡΑΜΜΑ 2).
Ένα από τα πιο σημαντικά ερωτήματα της σύγχρονης συναπτικής θεωρίας είναι ο λειτουργικός ρόλος της LTD. Αν ο λειτουργικός ρόλος της LTP στη μνημονική διαδικασία είναι η αύξηση της συναπτικής αποτελεσματικότητας και άρα η μείωση του αριθμού των συναπτικών σχέσεων που απαιτείται για να επέλθει συνειρμική συνενεργοποίηση ενός νευρωνικού υποπληθυσμού, ο ρόλος της LTD δεν είναι λιγότερο σημαντικός. Προτείνεται ότι η μείωση της συναπτικής αποτελεσματικότητας συγκεκριμένων νευρωνικών οδών είναι εντελώς απαραίτητη προκειμένου να είναι δυνατό να αναδυθούν λειτουργικά οι ενδυναμωμένες συνάψεις και οι νέες νευρωνικές οδοί που εγχαράχθηκαν με νέες πληροφορίες. Πράγματι, η λειτουργική επιτυχία της LTP φαίνεται να είναι αδύνατη χωρίς την LTD να εξασφαλίζει την απουσία του ηλεκτροκυτταρικού "θορύβου" ίσης έντασης από το ευρύτερο νευρωνικό πεδίο (background) που θα καθιστούσε αδύνατη τη διαυγή μνημονική λειτουργία. Η μνημονική εγχάραξη μιας αισθητηριακής εμπειρίας ή η μάθηση μιας διαδικασίας συμπεριφοράς δεν θα ήταν δυνατό να ολοκληρωθεί λειτουργικά σε παλιούς νευρώνες, οι οποίοι αποτελούν ήδη τμήματα παλαιότερων κυκλωμάτων, χωρίς τη μείωση της αποτελεσματικότητας συγκεκριμένων συναπτικών σχέσεων ώστε να ξεχωρίσουν οι νέες οδοί. Συνεπώς, ο λειτουργικός ρόλος της LTD δεν είναι η λήθη αλλά η δυνατότητα εγκαθίδρυσης της νέας μνημονικής εγχάραξης σε ένα παλαιό και ήδη πολυοδικό νευρωνικό κύκλωμα.
ΒΙΒΛΙΟΓΡΑΦΙΑ
Bliss TV & Collingridge GL. A synaptic model of long-term potentiation in the hippocampus. Nature, 1993. 361: 31-39.
Bliss TV & Lømo T. Neuroscience: Exploring the brain. [In: Bear MF, Connors BW, Paradiso MA. Neuroscience: Exploring the Brain. Williams & Wilkins, 1996. Baltimore, Maryland].
Brown TH, Chapman PF, Kairiss EW, Keenan CL. Long-term synaptic potentiation. Science, 1988.242: 724-728.
Chen Z, Sugimoto Y, Kamei C. Effects of intracere broven tricular injection of alpha-fluoromethylhistidine on radial maze performance in rats. Pharmacology and Biochemistry of Behavior, 1999, 64: 513-8.
Dudek SM. & Bear MF. Homosynaptic long-term depression in the area CA1 of the hippocampus and effects of N-methyl-D-aspartate receptor blocade. Proceeding of the National Academy of Sciences of the USA, 1992, 89: 4363-4367.
Kirkwood A & Bear MF. Elementary forms of synaptic plasticity in the visual cortex. Biological Research, 1995. 28: 73-80.
Kirkwood A, Rioult MC, Bear MF. Experience dependent modification of synaptic plasticity in visual cortex. Nature, 1996, 381: 526-528.
Levy WB & Steward O. Synapses as associative memory elements in the hippocampal formation. Brain Research, 1979, 175: 233-245.
Mayford M, Wang J, Kandel ER, O'Dell TJ. CaM-KII regulates the frequency-response function of hippocampal synapses for the production of both LTD and LTP.Cell,1995,81:891-904.
Petersen CC, Malenka RC, Nicoll RA, Hopfield JJ. All- or-none potentiation at CA3-CA1 synapses. Proceedings of the National Academy Of Sciences of the USA,1998,95:4732-4737.
Staubli VG & Zi ZX. The induction of homo- vs. heterosynaptic LTD in area CA1 of hippocampal slices from adult rats. Brain Research, 1996, 714: 169-176.
